遗传距离
衡量生物群体间遗传差异的指标。遗传距离可以用质量性状也可用数量性状来衡量。遗传距离的估计在探索品种起源、分析群体间亲缘关系、绘制系统发育树和预测杂种优势、指导亲本选配等方面有重要作用。估计的方法主要有Nei法、距离系数法、枢轴凝聚法和主成分法等。
Nei法 由奈(M.Nei)于1972年提出。它利用属性类别分布频率来计算生物群体间用质量性状衡量的遗传距离。计算公式为:
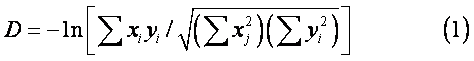
距离系数法 距离系数主要有以下几种:
Gower距离 第i个群体与第j个群体的Gower距离为:
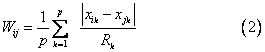
Canberra距离 第i个群体与第j个群体的Can-berra距离为:
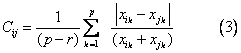
欧氏距离 第i个群体与第j个群体的欧氏距离为:
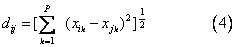
欧氏平方距离 即欧氏距离的平方
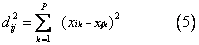
马氏距离 第i个群体与第j个群体的马氏距离为:
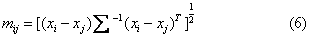
距离系数除以上五种外,还有绝对值距离、Bray-Curtis距离、弦距离、Minkowski距离、切比雪夫距离等, 它们或者受极端值影响大或者受性状相关的影响大, 应用较少。
上述利用观察值平均数计算的距离系数不能真正表示群体数量性状间的遗传距离。这是因为:①观察值平均数包含有环境因素的影响。②各性状间常常存在一定的相关性。③未采用方差分析法先对各性状是否存在遗传差异进行检验。针对这些问题,自20世纪50年代以来, 遗传育种科学家用多元统计方法陆续提出了估计群体数量性状遗传距离的枢轴凝聚法与主成分法。
枢轴凝聚法 由雷欧 (Rao) 于1952年提出, 其基本步骤是:
第一步 进行综合效应差异显著性检验。首先根据n个群体、p个性状的r次重复观测值对每 一性状进行方差分析,对每二个性状进行协方差分析,求出误差方差与协方差、群体方差与协方差,把误差方差与协方差与群体方差、协方差分别相加从而求出误差协方差矩阵W、误差+群体协方差矩阵S。
然后利用枢轴凝聚法求出误差协方差矩阵的枢轴凝聚元素Aii(i=1,2,…,p)、对数据进行转换的数值Gij(i,j=1, 2, … ,p); 求出误差+群体协方差矩阵的枢轴凝聚元素A′ii (i=1, 2, …, p)。
再求出误差协方差矩阵行列式W和误差+群体协方差矩阵的行列式|S|:

最后利用统计量V(stat.)进行综合效应差异显著性检验:
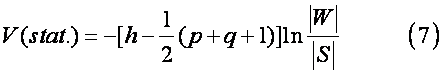
式中 h为误差+群体自由度;q=n-1为群体自由度。
因为V(stat.)近似服从自由度为pq的X2分布, 可对V(stat.)进行X2检验;若自由度pq>100,可转换成标准正态分布进行u检验:
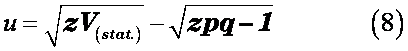
或者进行F检验:
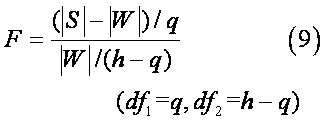
经X2检验或F检验显著,表明n个群体p个性状的综合效应间差异显著, 可进一步求各群体间的遗传距离。
第二步 对相关性状进行转换,计算遗传距离。转换的目的是把相关性状转换成非相关综合性状转换公式为:

式中 yij为转换后第i个群体第j个综合性状的平均数。
第i个群体与第j个群体间的遗传距离为:
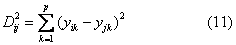
因为Dij2服从自由度为p的χ2分布,可对遗传距离D2ij的显著性进行χ2检验。
此法所求出的遗传距离进行了p个性状综合效应的差异显著性检验,克服了性状相关的影响,能较好地表示群体数量性状间的遗传差异,且可对遗传距离D2进行显著性检验。不足之处是包含有环境因素的影响。
利用遗传相关系数的主成分法 刘来福于1979年对此法作出了详细阐述,其基本步骤为:
第一步 对n个群体、p个性状、r次重复观测值进行方差分析,选留有显著遗传差异的性状(性状数仍记为p)。对选留的p个性状求出遗传方差σ2gj、遗传协方差σ2gij的估计值。在畜禽遗传育种研究中,对第j个性状进行方差分析时,常采用如下模型:
xijk=μj+gij+eijk (12)
(i=1,2,…,n;j=1,2,…,p;k=1,2,…,r)
式中 xijk为第i个群体第j个性状第k次观测值;μj为第j个性状的总平均数;gij为第i个群体第j个性状的基因型效应,服从N(0,σ2gj),eijk为随机误差,相互独立,且都服从N(0,σ2)。
在模型式(12)下,第j个性状的方差分析,第i、j两性状的方差分析一般形式为:
| 变异来源 | 自由度 | 均方 | 期望均方 | 均积 | 期望均积 |
| 群体 | n-1 | MSαj | σej2+rσgj2 | COVαij | σeij+rσgij |
| 误差 | n (r-1) | MSej | σej2 | COVeij | σeij |

